


News
-
人类造血干细胞可留存炎症应激记忆
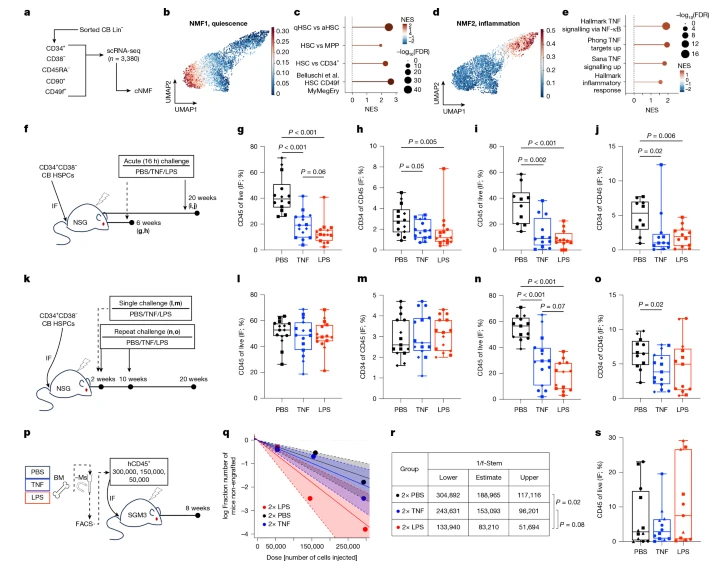 发表期刊:Nature|2026 年 5 月 27 日|开放获取 共同一作:Andy G. X. Zeng、Murtaza S. Nagree、Niels Asger Jakobsen 通讯共同负责人:Paresh Vyas、John E. Dick、Stephanie Z. Xie(谢昭雯) 摘要 炎症刺激可激活血细胞,进而诱发机体衰老与恶性肿瘤发生。造血干细胞(HSC)可在终生反复感染中存活、持续维系全生命周期造血功能,但人类造血干细胞如何应答并适应炎症应激,此前尚不明确。本研究构建人源造血干细胞异种移植炎症 - 恢复动物模型,结合单细胞多组学技术,解析炎症对人 HSC 的调控规律: 研究在人造血干细胞中发现两类转录组、表观遗传特征完全迥异的干细胞亚群;其中一类可留存既往炎症刺激的分子印记,被命名为炎症记忆造血干细胞(HSC-iM)。HSC-iM 长期维持静息状态、造血分化输出显著受抑。 在新冠肺炎康复患者、镰状细胞贫血患儿、自然衰老人群、克隆性造血(CH)患者的骨髓 / 外周造血样本中,HSC-iM 相关分子程序均显著富集,印证该细胞亚群在人体生理与疾病环境中真实存在。 携带克隆造血相关突变(DNMT3A/TET2)的 HSC-iM 可抵消炎症抑制效应、解除静息阻滞,促进干细胞活化与定向分化。 无论动物模型还是人体生理样本,HSC-iM 携带的促炎转录程序可向下传递至分化后的成熟免疫子代细胞。 人群队列分析证实:外周血细胞中 HSC-iM 特征基因富集度与全因死亡风险评分显著正相关,提示该新型造血干细胞亚群具备重要临床预后价值。 一、研究背景 人体造血系统需要持续海量血细胞供给,该需求由以 HSC 为顶端的层级化造血谱系实现。健康成人骨髓每秒可生成约 300 万个成熟血细胞,整个增殖分化过程被精密调控。人体内同步存在 5 万~20 万个功能性 HSC 持续参与造血。伴随年龄增长,HSC 功能逐步衰退、克隆多样性锐减,克隆造血发生率与血液肿瘤患病风险同步升高。人的一生反复遭遇细菌 / 病毒感染带来的急慢性炎症,但反复炎症如何改写 HSC 稳态、驱动衰老与造血克隆异常,分子机制不明。 既往小鼠实验证实:慢性炎症通过 TNF、NF-κB 通路活化 HSC,破坏干细胞自我更新与分化平衡,诱导表观修饰永久改变、加速干细胞老化,并促进携带致癌突变的造血克隆扩增。但小鼠结论无法直接套用人类:人类 HSC 存在高度异质性,不同干细胞亚群对炎症的应答是否一致、炎症记忆能否在干细胞中留存,长期是领域空白。 此外,癌前克隆造血突变常在确诊白血病数十年前即已发生,60 岁以上人群普遍检出低比例突变造血克隆,但仅极少数克隆最终恶性增殖。学界不清楚:炎症应激是否通过差异化改造 HSC 亚群,筛选、驱动突变克隆优势生长。 二、结果 1:稳态造血干细胞天然携带炎症预活化特征 研究首先利用脐带血来源长期造血干细胞(LT-HSC,免疫分型:Lin⁻CD34⁺CD38⁻CD45RA⁻CD90⁺CD49f⁺)开展单细胞转录组测序(3381 个细胞),通过非负矩阵分解(cNMF)解析细胞内在转录异质性,从中拆分出静息特征程序、炎症预应答程序两大固有基因模块;后续在 15590 个脐带血造血干 / 祖细胞单细胞多组学(RNA+ATAC)数据集重复验证:炎症程序伴随 AP-1、NF-κB 转录因子染色质开放特征,证明生理状态下部分 HSC 天然预先启动炎症相关染色质重塑,为后续炎症应答做好准备。 三、结果 2:构建人源化小鼠炎症模型,区分单次 / 反复炎症对 HSC 的差异化损伤 实验选用 NSG、NSG-SGM3、NSGW41 三类免疫缺陷小鼠,移植人脐带血 CD34⁺干祖细胞,分别使用细菌脂多糖 LPS(模拟脓毒症)、肿瘤坏死因子 TNF(衰老 / 重症新冠关键促炎因子)构建急性炎症、反复炎症两大模型: 单次炎症刺激:造模后短期(16h)人源骨髓植入水平、干祖细胞数量显著下降,但长期(18 周)可完全自我修复,干细胞重建能力无永久性损伤。 两次间隔反复炎症(移植后 2 周、10 周给药,恢复期 10 周):即便经过漫长恢复期,小鼠体内人源造血植入效率、CD34⁺祖细胞占比仍持续性下降;淋巴细胞谱系发生偏移:B 细胞占比下降、T 淋巴细胞比例显著升高;二次极限稀释移植证实:人类 HSC 在反复炎症后发生不可逆功能性重塑(区别于小鼠永久损伤),干细胞数量未减少,但分化潜能发生永久性偏移。 四、结果 3:首次发现 HSC-iM(炎症记忆造血干细胞)亚群 取反复炎症恢复期小鼠骨髓人源 HSPC 开展单细胞多组学(27492 个细胞),基于基因表达 + 染色质开放联合分群,将 HSC 拆分为HSC-I(常规造血干细胞)、HSC-II(后续定名 HSC-iM,炎症记忆干细胞)两个独立亚群: HSC-I:富集造血干性、巨核红系分化调控基因,以周期性活化、高效分化为特征,是机体日常造血主力; HSC-iM(原 HSC-II):NF-κB、AP-1、TGF-β 通路持续高开放,细胞长期锁定静息状态、基础造血输出显著降低;经 TNF/LPS 炎症刺激后,该亚群染色质与转录发生持久性修饰,即便炎症完全消退,数百种炎症相关转录因子(NFKB1、JUNB、HMGA1 等)仍持续高活性,永久留存过往炎症的分子记忆。 对比发现:HSC-iM 记忆程序和人体记忆 CD8⁺T 细胞表观调控高度同源(共用 AP-1 核心基序 TGAG/CTCA),但不同于卡介苗诱导的驯化免疫通路,证明HSC-iM 是独立于经典适应性免疫记忆、全新的干细胞记忆类型。 五、结果 4:人体真实疾病样本验证:HSC-iM 在炎症相关疾病、衰老中累积 研究跨多个人体原代骨髓数据库(合计 23048 例不同年龄段人 HSC),在四类临床样本验证 HSC-iM 生理存在: 重症新冠康复患者:ICU 收治后康复 2~4 个月患者骨髓 HSC 中,HSC-iM 特征基因、染色质开放程度远高于非新冠 ICU 对照与健康人;新冠特异性 HSC 特征基因可精准锚定 HSC-iM 亚群。 自然衰老:40~60 岁中年、60 岁以上老年受试者 HSC 中 HSC-iM 程序随年龄上调,研究者汇总全年龄段数据,筛选出 37 个衰老标志性基因,全部特异性富集于 HSC-iM,佐证慢性低度炎症(炎性衰老)持续诱导 HSC 向记忆型转化。 镰状细胞贫血(SCD)患儿:患儿骨髓 HSC 普遍富集 HSC-iM 特征,对应疾病终生慢性溶血诱发的持续性炎症,解释该病伴随的造血早衰表型。 六、结果 5:克隆造血(CH)突变优先改造 HSC-iM,驱动突变克隆扩增 研究依托DNMT3A/TET2突变克隆造血患者骨髓 TARGET-seq + 单细胞数据: 人体天然 HSC1 对应实验 HSC-I、HSC2 对应人体原生 HSC-iM;无论野生型还是携带 CH 突变的 HSC,均高度富集 HSC-iM 特征,有克隆造血人群体内 HSC-iM/HSC-I 比值显著高于健康人。 CH 突变(DNMT3A/TET2)仅在 HSC-iM 中引发大规模转录扰动,对 HSC-I 几乎无影响;突变可逆转 HSC-iM 固有的静息阻滞、解除分化抑制,使原本低输出的 HSC-iM 恢复造血分化能力,大量生成突变子代血细胞,最终推动突变克隆体积不断增大。 急性髓系白血病(AML)患者骨髓 HSC-iM 富集度显著升高,尤其DNMT3A/TET2突变亚型,建立「慢性炎症→HSC-iM 累积→CH 突变优势扩增→白血病易感」完整生物学链条。 七、结果 6:炎症记忆可跨代传递:HSC-iM 把促炎特征传给所有子代免疫细胞 通过供体天然基因多态性溯源克隆谱系,追踪 HSC 分化全链条: 源自 HSC-iM 的髓系祖细胞、单核细胞、淋巴细胞全程保留 HSC-iM 促炎转录特征,HSC-I 来源子代细胞则维持正常造血基因谱; 健康人、新冠康复患者骨髓单核细胞转录组,与其上游 HSC 的 HSC-iM 富集度高度相关,上游干细胞的炎症记忆直接决定下游成熟免疫细胞的先天炎症敏感性。 八、结果 7:HSC-iM 富集度关联人群全因死亡风险 依托加拿大安大略健康大队列(428 名受试者外周血单细胞测序),采用基于血常规的改良死亡风险评分(mIRS)分层: 年轻人群:外周 B 细胞、初始 T 细胞、树突状细胞内 HSC-iM 基因得分越高,个体全因死亡风险评分越高;HSC-I 富集则对应低死亡风险; 老年人群上述关联性减弱,提示幼年反复炎症塑造的 HSC-iM 储库,是决定终身远期健康与寿命的关键先天因素。 讨论 进化意义:HSC-iM 是人类造血系统在漫长进化中形成的适应性策略:遭遇反复感染后,部分 HSC 转入静息记忆状态,短期保护干细胞库免受炎症损伤;但长期代价是:HSC-iM 持续累积带来全身性慢性低度炎症、加速造血衰老,同时为致癌突变提供优势生存微环境。 临床价值:本研究首次证明人体造血系统存在干细胞层面炎症记忆,解释了「既往重症感染(新冠、脓毒症)多年后仍提升慢性病、血液病患病风险」的临床现象;未来可将 HSC-iM 分子特征作为新型生物标志物,用于衰老分层、克隆造血筛查、远期死亡风险预判。 待解决问题:目前缺少 HSC-iM 特异性分选标志物,无法活体分离该亚群;HSC-iM 能否通过药物干预逆转表观记忆、改善炎性衰老与克隆造血,是后续转化研究核心方向。 实验方法精简译 样本来源:人脐带血获加拿大三家医院伦理审批;人体骨髓样本来自髋置换手术废弃组织、安大略健康队列,全部受试者签署知情同意(伦理号:UHN REB# 02-0763)。 动物实验:NSG/NSG-SGM3 小鼠 8~12 周龄,经辐照后股骨原位移植 8000~10000 个 CD34⁺CD38⁻细胞;TNF(5μg / 只)、LPS(40μg / 只)腹腔给药构建炎症模型。 测序技术:10×Genomics 单细胞多组学(RNA+ATAC)、TARGET-seq(单细胞转录 + 定点突变分型);数据分析采用 Seurat、Signac、SCENIC+、AUCell、GSEA 等生信算法。 数据存储:测序原始数据存 GEO(GSE249479)、欧洲基因组档案 EGA;分析代码开源于 Zenodo。 核心名词汇总 英文缩写 中文译名 HSC 造血干细胞 LT-HSC 长期造血干细胞 HSC-iM 炎症记忆造血干细胞 CH 克隆性造血 LPS 脂多糖(内毒素) TNF 肿瘤坏死因子 α scMultiome 单细胞多组学(转录 + 染色质可及性)More
发表期刊:Nature|2026 年 5 月 27 日|开放获取 共同一作:Andy G. X. Zeng、Murtaza S. Nagree、Niels Asger Jakobsen 通讯共同负责人:Paresh Vyas、John E. Dick、Stephanie Z. Xie(谢昭雯) 摘要 炎症刺激可激活血细胞,进而诱发机体衰老与恶性肿瘤发生。造血干细胞(HSC)可在终生反复感染中存活、持续维系全生命周期造血功能,但人类造血干细胞如何应答并适应炎症应激,此前尚不明确。本研究构建人源造血干细胞异种移植炎症 - 恢复动物模型,结合单细胞多组学技术,解析炎症对人 HSC 的调控规律: 研究在人造血干细胞中发现两类转录组、表观遗传特征完全迥异的干细胞亚群;其中一类可留存既往炎症刺激的分子印记,被命名为炎症记忆造血干细胞(HSC-iM)。HSC-iM 长期维持静息状态、造血分化输出显著受抑。 在新冠肺炎康复患者、镰状细胞贫血患儿、自然衰老人群、克隆性造血(CH)患者的骨髓 / 外周造血样本中,HSC-iM 相关分子程序均显著富集,印证该细胞亚群在人体生理与疾病环境中真实存在。 携带克隆造血相关突变(DNMT3A/TET2)的 HSC-iM 可抵消炎症抑制效应、解除静息阻滞,促进干细胞活化与定向分化。 无论动物模型还是人体生理样本,HSC-iM 携带的促炎转录程序可向下传递至分化后的成熟免疫子代细胞。 人群队列分析证实:外周血细胞中 HSC-iM 特征基因富集度与全因死亡风险评分显著正相关,提示该新型造血干细胞亚群具备重要临床预后价值。 一、研究背景 人体造血系统需要持续海量血细胞供给,该需求由以 HSC 为顶端的层级化造血谱系实现。健康成人骨髓每秒可生成约 300 万个成熟血细胞,整个增殖分化过程被精密调控。人体内同步存在 5 万~20 万个功能性 HSC 持续参与造血。伴随年龄增长,HSC 功能逐步衰退、克隆多样性锐减,克隆造血发生率与血液肿瘤患病风险同步升高。人的一生反复遭遇细菌 / 病毒感染带来的急慢性炎症,但反复炎症如何改写 HSC 稳态、驱动衰老与造血克隆异常,分子机制不明。 既往小鼠实验证实:慢性炎症通过 TNF、NF-κB 通路活化 HSC,破坏干细胞自我更新与分化平衡,诱导表观修饰永久改变、加速干细胞老化,并促进携带致癌突变的造血克隆扩增。但小鼠结论无法直接套用人类:人类 HSC 存在高度异质性,不同干细胞亚群对炎症的应答是否一致、炎症记忆能否在干细胞中留存,长期是领域空白。 此外,癌前克隆造血突变常在确诊白血病数十年前即已发生,60 岁以上人群普遍检出低比例突变造血克隆,但仅极少数克隆最终恶性增殖。学界不清楚:炎症应激是否通过差异化改造 HSC 亚群,筛选、驱动突变克隆优势生长。 二、结果 1:稳态造血干细胞天然携带炎症预活化特征 研究首先利用脐带血来源长期造血干细胞(LT-HSC,免疫分型:Lin⁻CD34⁺CD38⁻CD45RA⁻CD90⁺CD49f⁺)开展单细胞转录组测序(3381 个细胞),通过非负矩阵分解(cNMF)解析细胞内在转录异质性,从中拆分出静息特征程序、炎症预应答程序两大固有基因模块;后续在 15590 个脐带血造血干 / 祖细胞单细胞多组学(RNA+ATAC)数据集重复验证:炎症程序伴随 AP-1、NF-κB 转录因子染色质开放特征,证明生理状态下部分 HSC 天然预先启动炎症相关染色质重塑,为后续炎症应答做好准备。 三、结果 2:构建人源化小鼠炎症模型,区分单次 / 反复炎症对 HSC 的差异化损伤 实验选用 NSG、NSG-SGM3、NSGW41 三类免疫缺陷小鼠,移植人脐带血 CD34⁺干祖细胞,分别使用细菌脂多糖 LPS(模拟脓毒症)、肿瘤坏死因子 TNF(衰老 / 重症新冠关键促炎因子)构建急性炎症、反复炎症两大模型: 单次炎症刺激:造模后短期(16h)人源骨髓植入水平、干祖细胞数量显著下降,但长期(18 周)可完全自我修复,干细胞重建能力无永久性损伤。 两次间隔反复炎症(移植后 2 周、10 周给药,恢复期 10 周):即便经过漫长恢复期,小鼠体内人源造血植入效率、CD34⁺祖细胞占比仍持续性下降;淋巴细胞谱系发生偏移:B 细胞占比下降、T 淋巴细胞比例显著升高;二次极限稀释移植证实:人类 HSC 在反复炎症后发生不可逆功能性重塑(区别于小鼠永久损伤),干细胞数量未减少,但分化潜能发生永久性偏移。 四、结果 3:首次发现 HSC-iM(炎症记忆造血干细胞)亚群 取反复炎症恢复期小鼠骨髓人源 HSPC 开展单细胞多组学(27492 个细胞),基于基因表达 + 染色质开放联合分群,将 HSC 拆分为HSC-I(常规造血干细胞)、HSC-II(后续定名 HSC-iM,炎症记忆干细胞)两个独立亚群: HSC-I:富集造血干性、巨核红系分化调控基因,以周期性活化、高效分化为特征,是机体日常造血主力; HSC-iM(原 HSC-II):NF-κB、AP-1、TGF-β 通路持续高开放,细胞长期锁定静息状态、基础造血输出显著降低;经 TNF/LPS 炎症刺激后,该亚群染色质与转录发生持久性修饰,即便炎症完全消退,数百种炎症相关转录因子(NFKB1、JUNB、HMGA1 等)仍持续高活性,永久留存过往炎症的分子记忆。 对比发现:HSC-iM 记忆程序和人体记忆 CD8⁺T 细胞表观调控高度同源(共用 AP-1 核心基序 TGAG/CTCA),但不同于卡介苗诱导的驯化免疫通路,证明HSC-iM 是独立于经典适应性免疫记忆、全新的干细胞记忆类型。 五、结果 4:人体真实疾病样本验证:HSC-iM 在炎症相关疾病、衰老中累积 研究跨多个人体原代骨髓数据库(合计 23048 例不同年龄段人 HSC),在四类临床样本验证 HSC-iM 生理存在: 重症新冠康复患者:ICU 收治后康复 2~4 个月患者骨髓 HSC 中,HSC-iM 特征基因、染色质开放程度远高于非新冠 ICU 对照与健康人;新冠特异性 HSC 特征基因可精准锚定 HSC-iM 亚群。 自然衰老:40~60 岁中年、60 岁以上老年受试者 HSC 中 HSC-iM 程序随年龄上调,研究者汇总全年龄段数据,筛选出 37 个衰老标志性基因,全部特异性富集于 HSC-iM,佐证慢性低度炎症(炎性衰老)持续诱导 HSC 向记忆型转化。 镰状细胞贫血(SCD)患儿:患儿骨髓 HSC 普遍富集 HSC-iM 特征,对应疾病终生慢性溶血诱发的持续性炎症,解释该病伴随的造血早衰表型。 六、结果 5:克隆造血(CH)突变优先改造 HSC-iM,驱动突变克隆扩增 研究依托DNMT3A/TET2突变克隆造血患者骨髓 TARGET-seq + 单细胞数据: 人体天然 HSC1 对应实验 HSC-I、HSC2 对应人体原生 HSC-iM;无论野生型还是携带 CH 突变的 HSC,均高度富集 HSC-iM 特征,有克隆造血人群体内 HSC-iM/HSC-I 比值显著高于健康人。 CH 突变(DNMT3A/TET2)仅在 HSC-iM 中引发大规模转录扰动,对 HSC-I 几乎无影响;突变可逆转 HSC-iM 固有的静息阻滞、解除分化抑制,使原本低输出的 HSC-iM 恢复造血分化能力,大量生成突变子代血细胞,最终推动突变克隆体积不断增大。 急性髓系白血病(AML)患者骨髓 HSC-iM 富集度显著升高,尤其DNMT3A/TET2突变亚型,建立「慢性炎症→HSC-iM 累积→CH 突变优势扩增→白血病易感」完整生物学链条。 七、结果 6:炎症记忆可跨代传递:HSC-iM 把促炎特征传给所有子代免疫细胞 通过供体天然基因多态性溯源克隆谱系,追踪 HSC 分化全链条: 源自 HSC-iM 的髓系祖细胞、单核细胞、淋巴细胞全程保留 HSC-iM 促炎转录特征,HSC-I 来源子代细胞则维持正常造血基因谱; 健康人、新冠康复患者骨髓单核细胞转录组,与其上游 HSC 的 HSC-iM 富集度高度相关,上游干细胞的炎症记忆直接决定下游成熟免疫细胞的先天炎症敏感性。 八、结果 7:HSC-iM 富集度关联人群全因死亡风险 依托加拿大安大略健康大队列(428 名受试者外周血单细胞测序),采用基于血常规的改良死亡风险评分(mIRS)分层: 年轻人群:外周 B 细胞、初始 T 细胞、树突状细胞内 HSC-iM 基因得分越高,个体全因死亡风险评分越高;HSC-I 富集则对应低死亡风险; 老年人群上述关联性减弱,提示幼年反复炎症塑造的 HSC-iM 储库,是决定终身远期健康与寿命的关键先天因素。 讨论 进化意义:HSC-iM 是人类造血系统在漫长进化中形成的适应性策略:遭遇反复感染后,部分 HSC 转入静息记忆状态,短期保护干细胞库免受炎症损伤;但长期代价是:HSC-iM 持续累积带来全身性慢性低度炎症、加速造血衰老,同时为致癌突变提供优势生存微环境。 临床价值:本研究首次证明人体造血系统存在干细胞层面炎症记忆,解释了「既往重症感染(新冠、脓毒症)多年后仍提升慢性病、血液病患病风险」的临床现象;未来可将 HSC-iM 分子特征作为新型生物标志物,用于衰老分层、克隆造血筛查、远期死亡风险预判。 待解决问题:目前缺少 HSC-iM 特异性分选标志物,无法活体分离该亚群;HSC-iM 能否通过药物干预逆转表观记忆、改善炎性衰老与克隆造血,是后续转化研究核心方向。 实验方法精简译 样本来源:人脐带血获加拿大三家医院伦理审批;人体骨髓样本来自髋置换手术废弃组织、安大略健康队列,全部受试者签署知情同意(伦理号:UHN REB# 02-0763)。 动物实验:NSG/NSG-SGM3 小鼠 8~12 周龄,经辐照后股骨原位移植 8000~10000 个 CD34⁺CD38⁻细胞;TNF(5μg / 只)、LPS(40μg / 只)腹腔给药构建炎症模型。 测序技术:10×Genomics 单细胞多组学(RNA+ATAC)、TARGET-seq(单细胞转录 + 定点突变分型);数据分析采用 Seurat、Signac、SCENIC+、AUCell、GSEA 等生信算法。 数据存储:测序原始数据存 GEO(GSE249479)、欧洲基因组档案 EGA;分析代码开源于 Zenodo。 核心名词汇总 英文缩写 中文译名 HSC 造血干细胞 LT-HSC 长期造血干细胞 HSC-iM 炎症记忆造血干细胞 CH 克隆性造血 LPS 脂多糖(内毒素) TNF 肿瘤坏死因子 α scMultiome 单细胞多组学(转录 + 染色质可及性)More -
不同转录因子调控婴儿与成人记忆 T 细胞的组织适应性及效应功能
 不同转录因子调控婴儿与成人记忆 T 细胞的组织适应性及效应功能 期刊:Nature Immunology(2026) 发表时间:2026 年 6 月 2 日 DOI:10.1038/s41590-026-02535-1 摘要 生命早期是建立记忆 T 细胞库的关键阶段,记忆 T 细胞在婴儿期可快速定植于黏膜组织,但这些新生记忆 T 细胞的免疫保护能力弱于成人。本研究对婴儿(2–9 月龄)与成人(40–63 岁)器官供体的淋巴组织与黏膜组织中静息及 CD3+CD28 抗体活化的 T 细胞进行单细胞 RNA 测序,探究人类记忆 T 细胞功能调控的年龄依赖性机制。 结果显示,婴儿CCL5 + 效应记忆 T 细胞的效应功能弱于成人。转录因子调控网络分析鉴定出HELIOS与KLF6分别为婴儿与成人组织记忆 T 细胞状态的核心调控因子。结合单细胞核 RNA 测序、转座酶可及染色质测序与 CRISPR–Cas9 敲除实验,证实HELIOS(IKZF2)是婴儿 CCL5 + 效应记忆 T 细胞特异性转录程序的关键调控因子,可抑制 SELL+CCR7 + 初始 T 细胞 / 中央记忆 T 细胞的效应功能。本研究揭示了生命早期调控 T 细胞功能状态的核心分子机制。 正文 一、引言 生命早期适应性免疫系统的成熟对建立终身免疫记忆与防御能力至关重要。婴幼儿阶段会持续接触共生菌、致病菌、疫苗及环境抗原,触发强烈的初次免疫应答并形成记忆 T 细胞,介导后续的回忆反应。但相较于成人,婴儿对呼吸道、胃肠道等常见病原体高度易感,提示其记忆应答发育尚不完善。 小鼠研究已证实,组织驻留记忆 T 细胞(TRM)介导黏膜与屏障部位的免疫保护;但人类生命早期组织记忆应答的功能与建立机制仍不明确,而这对优化婴幼儿疫苗策略与免疫治疗至关重要。 婴儿小鼠与人类的 T 细胞以初始 T 细胞(TN)为主。婴儿小鼠感染模型显示,婴儿 TN可分化为效应 T 细胞(Teff),但针对全身性与呼吸道病毒感染的循环及组织定位记忆 T 细胞生成量更少、保护力更弱;婴儿疫苗诱导的记忆 T 细胞数量与保护力也显著低于成年小鼠。 婴儿与成人 T 细胞的原发性应答差异源于细胞固有机制,包括 T 细胞受体(TCR)敏感性、造血前体细胞特性及胸腺新近输出细胞的影响。然而,婴儿记忆 T 细胞应答欠佳的分子机制在小鼠中尚未阐明,在人类中更是完全未知。 借助人类跨年龄段器官供体的多组织样本,研究者已明确人类组织驻留记忆 T 细胞(TRM)的核心特征、黏膜组织分区、位点特异性适应及年龄稳定性。出生后 1–2 年内,具有经典 CD45RA−CCR7−效应记忆 T 细胞(TEM)表型并表达 CD69(TRM标志性分子)的 T 细胞迅速定植于黏膜组织,但这类细胞的核心及组织特异性 TRM标志物表达水平更低,且产生促炎细胞因子(IFN-γ、TNF)的能力弱于成人及年长儿童的黏膜 TRM。目前尚不清楚调控婴儿记忆 T 细胞独特表型的转录机制,以及组织适应性黏膜记忆 T 细胞的成熟通路。 本研究采用单细胞 RNA 测序(scRNA‑seq)分析婴儿与成人淋巴组织、黏膜组织来源的 T 细胞(含 CD3+CD28 抗体活化组),应用一致性单层级泊松因子分解(consensus‑scHPF)解析婴儿与成人 T 细胞活化、效应功能及组织适应相关的转录状态。 研究发现,婴儿组织记忆 T 细胞呈现干细胞样转录特征(TCF1、LEF1、SOX4),但对 TCR 介导刺激的转录应答弱于成人。通过概率建模与调控网络推断,鉴定出HELIOS(IKZF2)与KLF6为年龄特异性转录因子,分别调控婴儿与成人 T 细胞的功能与组织适应。利用 CRISPR–Cas9 敲除、单细胞核 RNA 测序(snRNA‑seq)与转座酶可及染色质测序(ATAC‑seq)证实,HELIOS 是婴儿记忆 T 细胞的关键调控因子,可维持干细胞样状态、抑制效应分化及组织特征表达。综上,本研究揭示年龄调控记忆 T 细胞命运与功能的核心机制,为生命早期 T 细胞应答靶向干预提供理论依据。 二、结果 2.1 单细胞 RNA 测序绘制不同年龄与组织的 T 细胞亚群图谱 为明确生命早期不同组织 T 细胞的转录编程,本研究对婴儿(n=4,2–9 月龄,男女各 2 例)与成人(n=4,40–63 岁,男性 4 例)的淋巴组织、黏膜组织 T 细胞进行 scRNA‑seq 检测。样本取自离世器官捐献者,包括淋巴器官(骨髓、脾脏、扁桃体、肠道派尔集合淋巴结、肺 / 空肠 / 结肠相关淋巴结)与黏膜 / 屏障组织(肺、空肠、回肠、结肠)。 经磁珠阴性分选富集 T 细胞,分为静息组(培养基培养)与活化组(CD3+CD28 抗体刺激),采用 10x Genomics Chromium 平台进行单细胞测序。数据集与已发表的成人器官供体及健康献血者静息 / 活化 T 细胞数据合并,最终获得约 27.5 万个跨 12 种组织的 T 细胞单细胞图谱。 合并数据集采用朴素贝叶斯分类器划分 T 细胞亚群:SELL+CCR7 + 初始 T 细胞 / 中央记忆 T 细胞(TN/TCM)、CCL5 + 效应记忆 T 细胞(TEM,含 CD4 + 与 CD8+)、CD4+FOXP3 + 调节性 T 细胞(Treg)及 γδ T 细胞。均匀流形逼近与投影(UMAP)显示,年龄与刺激条件是导致转录组变异的主要因素。婴儿与成人各 T 细胞亚群的经典标志物表达高度保守。 为对比婴儿与成人跨组织 T 细胞谱,聚焦两组共有的组织样本:血液、骨髓、肺 / 空肠相关淋巴结、脾脏、肺、空肠、结肠。婴儿血液与淋巴部位的 T 细胞以 SELL+CCR7+ TN/TCM为主,仅少量 CD4+FOXP3+ Treg及 CCL5+ TEM/γδ T 细胞;婴儿黏膜组织与脾脏中 CCL5+ TEM比例更高,尤以 CD8+CCL5+ TEM显著,肠道 T 细胞几乎均为 CCL5+ TEM,与既往报道一致。 与之相反,成人黏膜组织与脾脏中 CCL5+ TEM占绝对优势,比例显著高于 SELL+CCR7+ TN/TCM;成人多数组织中 CD8+ SELL+CCR7+ TN/TCM比例远低于 CD4 + 亚群。上述结果表明,婴儿与成人 T 细胞亚群组成存在组织特异性年龄差异,与既往流式细胞术结果相符。 2.2 婴儿效应记忆 T 细胞对 TCR 刺激呈低反应性 婴儿与成人 T 细胞亚群的差异表达分析显示,CCL5+ TEM在静息与活化状态下均存在显著年龄差异。静息状态下,婴儿 CCL5+ TEM高表达干细胞样记忆相关转录因子TCF7(TCF1)、LEF1、KLF2,以及 Wnt 信号通路 TCF‑LEF 家族辅因子SOX4与成人 Treg特征性分子IKZF2(HELIOS)。 婴儿 CD4+CCL5+ TEM富集 Th2 驱动转录因子GATA3,CD8 + 亚群高表达组织驻留相关ZNF683(Hobit)与干细胞样记忆前体标志物ID3。而成人 CCL5+ TEM偏好表达组织适应与黏附相关基因,如LGALS1、LGALS3、ANXA1、VIM、ITGA1。 流式细胞术证实,婴儿脾脏 CD8+CCR7−TEM中 TCF1 与 LEF1 表达显著高于成人,CD4+CCR7−TEM中仅 LEF1 显著升高。 差异表达分析发现,成人 CD3+CD28 抗体刺激 T 细胞(尤其 CCL5+ TEM)中促炎细胞因子、趋化因子与细胞毒性介质(IFNG、IL‑2、TNF、CCL3、CCL4、GZMB、PRF1)的转录本诱导更强。流式细胞术显示,成人脾脏 CD4+CCR7−TEM与 CD8+CCR7−TEM中 IFNγ、IL‑2、TNF 阳性细胞比例显著高于婴儿。 综上,婴儿 CCL5+ TEM呈现独特转录状态:干性相关转录因子高表达,效应分子表达低于成人 CCL5+ TEM。 2.3 T 细胞呈现年龄与组织特异性表达程序 为揭示婴儿与成人组织 T 细胞的独特基因表达程序,采用consensus‑scHPF(贝叶斯因子分解法)挖掘单细胞中共同表达的潜在转录特征(因子),并以年龄、组织定位、亚群、谱系、活化状态为协变量进行多元线性回归,关联各因子与临床表型。 分析鉴定出 18 种独立特征,对应 T 细胞亚群、代谢、组织适应、活化状态及 / 或效应功能。 consensus‑scHPF 揭示 3 种与组织定位及适应相关、且婴儿与成人存在差异的因子: 婴儿组织因子:在婴儿 CCL5+ TEM中高差异表达,含 SMC4、NCR3、CXXC5、LAYN、IKZF2,各组织中婴儿 CCL5+ TEM均显著富集。 组织特征因子:在成人 CD4+/CD8+CCL5+ TEM中富集,含 S100A4/6、CRIP1、LGALS1、KLRB1、ANXA2。 肠道驻留因子:以肠道归巢与黏附标志物(CCR9、ITGA1、CTNNA1)及组织驻留记忆 T 细胞发育相关因子(AHR、JUN、FOSB)为特征,显著偏向成人 CD4+/CD8+CCL5+ TEM(尤其黏膜组织)。 另有 3 种与效应状态相关的因子:细胞毒性因子(GZMK、GNLY、GZMA、NKG7、PRF1)、趋化 / 细胞毒性因子(GZMB、GZMH、FASLG、PRF1、CCL3、CCL4)、炎性细胞因子因子(IFNG、IL‑2、TNF、CSF2、LTA)。三者均在成人 CCL5+ TEM中富集,且后两者在活化 CCL5+ TEM中高于静息组。 整体表明,婴儿与成人 T 细胞的组织适应程序及TCR 刺激应答存在显著差异。 对 CD3+CD28 抗体刺激后 CCL5+ TEM的分析显示,成人活化 CCL5+ TEM(尤其 CD8 + 亚群)中炎性细胞因子与趋化 / 细胞毒性因子的核心基因表达更高。扩散映射活化轨迹显示,成人 CD8+CCL5+ TEM沿活化轴的细胞数量远多于婴儿,提示婴儿 T 细胞刺激后的转录应答减弱、效应功能受限。 在 3 种组织驻留 / 适应相关因子中,婴儿组织因子核心基因高度富集于婴儿 CD4+/CD8+CCL5+ TEM;组织特征因子在婴儿与成人 CCL5+ TEM中广泛表达;肠道驻留因子仅富集于成人肠道部位(空肠、结肠、空肠相关淋巴结)的 CCL5+ TEM。空肠静息 CCL5+ TEM的成熟轨迹显示,婴儿组织因子与肠道驻留因子分别特异性富集于婴儿与成人细胞,而组织特征因子为两者共有,提示其代表空肠组织适应的中间状态。 2.4 T 细胞激活年龄特异性转录因子 为鉴定执行组织与年龄特异性表达程序的转录因子,应用ARACNe 算法重构基因调控网络,生成各转录因子的靶基因集(调控子);再通过基因集富集分析(GSEA)将转录因子与 scHPF 因子定义的细胞状态关联。 该分析鉴定出多种已知转录因子‑T 细胞状态关系:IRF1 与 STAT1 调控干扰素信号应答、NFKB1 调控活化、IRF7 调控 Treg干扰素应答、ZEB2 调控细胞毒性 T 细胞功能、KLF2/TCF1/LEF1 调控初始 T 细胞干性与静息态。同时发现,HELIOS(IKZF2)是婴儿组织因子的首要调控因子,KLF6是组织特征因子与肠道驻留因子的首要调控因子。 进一步通过VIPER 分析在单细胞水平推断转录因子活性。空肠组织适应轨迹的映射结果证实:HELIOS 活性仅存在于婴儿,KLF6 活性随婴儿向成人过渡而增强,并在所有成人 CD4+/CD8+CCL5+ TEM中升高。 组织驻留相关转录因子中,RUNX3 在婴儿与成人 CCL5+ TEM均有活性,AHR 活性显著富集于成人 CCL5+ TEM;干性转录因子 LEF1、TCF1 活性显著富集于婴儿 CCL5+ TEM。LEF1 调控子与组织特征因子、肠道驻留因子核心基因(含 KLF6)存在大量抑制性相互作用,提示 LEF1 可能拮抗组织适应所需的转录程序。 综上,本分析发现与 T 细胞状态相关的关键转录因子,并确定HELIOS是 T 细胞中婴儿特异性组织适应程序的候选调控因子。 2.5 HELIOS 调控婴儿 T 细胞的效应功能 为探究 HELIOS 在婴儿 CD8+ TEM中的表达与活性调控,对额外 3 例婴儿(0.25–1 岁,男性)与 2 例成人(45–51 岁,男性)供体的脾细胞进行snRNA‑seq/ATAC‑seq 联合测序。通过婴儿与成人 T 细胞的染色质可及性差异,鉴定年龄依赖性基因表达的顺式调控元件,并以差异可及染色质峰中的转录因子基序富集作为年龄依赖性转录因子活性的正交验证。 整合 snRNA‑seq/ATAC‑seq 的 CD8+CCL5+ TEM图谱,鉴定出 1 种婴儿特异性细胞状态与 2 种成人细胞状态。构建基于 mRNA 的拟时轨迹,解析从婴儿状态向两种成人状态过渡过程中的基因表达与转录因子活性变化。 第一种成人 CD8+CCL5+ TEM状态:细胞毒性与活化标志物(GZMA、GZMH、GNLY、CCL4、CCL5)高于婴儿状态; 第二种成人 CD8+CCL5+ TEM状态:共刺激标志物(CD28、ICOS)与转录因子 BATF 高表达。 结合同一细胞核的基因表达与染色质可及性数据,进行转录因子基序富集分析,并沿婴儿→成人拟时轨迹映射转录因子活性。结果显示,HELIOS 活性在两种轨迹中均显著富集于婴儿 CD8+CCL5+ TEM状态,在独立器官供体队列中从染色质水平独立验证了 scRNA‑seq 与调控网络的发现。 利用 CD8+CCL5+ TEM的 snATAC‑seq 数据,评估 IKZF2 表达是否受年龄依赖性顺式调控。在 IKZF2 位点上游鉴定出 1 个顺式调控元件,其染色质可及性峰在婴儿 CD8+CCL5+ TEM中富集;该区域含多个婴儿特异性活性转录因子(SOX4、ETV1、TCF7L1)的结合基序,以及人群遗传研究中与免疫细胞 IKZF2 表达相关的表达数量性状位点(eQTLs),提示 IKZF2 表达通过该顺式调控元件受成人 CD8+CCL5+ TEM中活性减弱的转录因子调控。 流式细胞术显示,婴儿 T 细胞中 HELIOS 表达显著高于成人,涵盖 CD8+CD45RA+CCR7+ TN、CD8+CD45RA−CCR7+ TCM、CD8+CCR7−TEM,CD4+CD45RA+CCR7+ TN中差异稍弱。婴儿与成人的 CD8+/CD4+ T 细胞经 CD3+CD28 抗体刺激后,HELIOS 的 mRNA 与蛋白表达均未上调。 采用CRISPR–Cas9 基因编辑敲除原代婴儿 T 细胞中的 HELIOS。对 2 例婴儿供体(2–3 月龄,男性)脾脏分离的 CD3+ T 细胞,转染靶向 IKZF2 的 Cas9 核糖核蛋白(未预先刺激 T 细胞),流式细胞术检测敲除效率。结果显示,HELIOS 敲除使 γδTCR−FOXP3−CD3+ T 细胞中 HELIOS + 细胞比例降低约 60%。 对静息状态下婴儿 HELIOS 敲除与野生型 CD3+ T 细胞的 scRNA‑seq 分析显示,敲除组 CD8+CCL5+ TEM中 HELIOS 激活靶基因(LAYN、PDE7B、CHST2、SMC4、CXXC5)表达下调,CRISPR–Cas9 敲除验证了计算推断的 HELIOS 转录靶标。 HELIOS 敲除与野生型 T 细胞的大部分转录差异出现在 CD3+CD28 抗体刺激的 CD4+SELL+CCR7+ TN–TCM与 CD8+SELL+CCR7+ TN–TCM中,包括促炎细胞因子、趋化因子(IL‑2、CSF2、XCL1、XCL2、CCL3、CCL4、CCL20)与共刺激分子(TIGIT、KLRB1、TNFRSF4、TNFRSF9、TNFRSF18、TNFSF14、FCER1G、IL18R1、IL1R1)转录本表达升高,提示TCR 介导的活化增强。 4 小时 PMA + 离子霉素刺激后,婴儿 HELIOS 敲除组 CD4+CD45RA−CCR7+ TCM与 CD4+CCR7−TEM中 IL‑2 + 细胞比例高于野生型。scHPF 分析显示,与 T 细胞活化及效应功能相关的趋化 / 细胞毒性因子、炎性细胞因子因子,在 HELIOS 敲除婴儿 T 细胞(尤其 CD4+/CD8+SELL+CCR7+ TN–TCM)中显著富集,表明HELIOS 缺失使婴儿 T 细胞对刺激的转录应答更接近成人。 静息状态下 HELIOS 敲除 T 细胞的转录因子分析显示,KLF6及其他 T 细胞效应功能调控因子(IRF1、IRF2、STAT5A、IKZF3、RORA)表达升高,多数与成人 SELL+CCR7+ TN–TCM相关。CD3+CD28 抗体刺激后,婴儿 HELIOS 敲除 SELL+CCR7+ TN–TCM中干性转录因子 TCF7、LEF1 表达降低(二者为婴儿野生型 SELL+CCR7+ TN–TCM的特征)。 综上数据表明,HELIOS 缺失驱动婴儿 T 细胞从婴儿样转录状态向成人样状态转变。 三、讨论 本研究系统分析了人类组织中婴儿与成人 T 细胞的功能应答及组织定位,发现婴儿期记忆 T 细胞主要局限于黏膜部位,功能呈低反应性,组织驻留关键分子表达低于成人记忆 T 细胞;同时鉴定出HELIOS与KLF6分别为调控婴儿与成人组织记忆 T 细胞功能及组织驻留的核心转录因子。 snATAC‑seq 证实 HELIOS 调控活性特异性富集于婴儿记忆 T 细胞;婴儿 T 细胞中 HELIOS 敲除可增强活化与效应功能,使其趋近成人记忆 T 细胞。本研究揭示了控制组织适应与效应应答的年龄特异性转录程序,作为生命早期限制记忆 T 细胞功能的核心机制。 通过 scRNA‑seq 对比黏膜与淋巴部位静息、活化状态下婴儿与成人 T 细胞的转录谱,从多方面证实婴儿 TEM处于分化程度更低、功能未成熟的状态: 婴儿 TEM即使在刺激后仍维持干性标志物(SOX4、TCF7、LEF1)表达,成人 TEM则不表达; 婴儿 TEM的组织驻留标志物(ITGA1、ITGAE、CXCR6)与位点特异性适应标志物(如 CCR9)表达低于成人,与既往报道一致; 既往研究显示婴儿 TEM经 PMA + 离子霉素非特异性活化后,效应细胞因子分泌少于成人 TEM;本研究进一步发现,婴儿记忆 T 细胞对TCR 偶联刺激基本无反应,不具备成人 TEM活化标志性的快速回忆应答,提示存在婴儿 T 细胞固有程序介导的耐受状态。 本研究将HELIOS鉴定为婴儿 T 细胞活化与分化的核心转录调控因子与抑制因子。婴儿 T 细胞中 HELIOS 转录因子基序活性显著高于成人;特异性敲除 HELIOS 可增强婴儿 T 细胞的活化与效应功能。值得注意的是,静息婴儿 T 细胞中敲除 HELIOS 可诱导成人样转录因子(含 KLF6)表达,CD3+CD28 抗体刺激后高表达 CCL3、CCL4、IL‑2 等趋化因子与细胞因子基因,趋近成人组织记忆 T 细胞。 HELIOS 最初被发现表达于小鼠与人 Treg,可沉默 IL2 位点;IKZF2 功能缺失与错义突变患者的 T 细胞呈现 IL‑2 分泌增强或异常。外周血新生儿与胎儿 T 细胞(以初始表型为主)的 HELIOS 表达也高于成人。本研究发现,婴儿初始 T 细胞的 IKZF2 表达高于成人,且婴儿 TEM的 IKZF2 表达显著高于初始 T 细胞,据此提出HELIOS 作为组织定位 TEM中促炎应答与驻留的细胞内源性抑制因子。 通过基因调控网络推断,将KLF6鉴定为跨多组织的组织特征及肠道特异性驻留特征的候选转录调控因子,与成人空肠 T 细胞的肠道驻留特征相似。小鼠研究中 Klf6 被鉴定为 TRM特异性转录因子,在抗原特异性 T 细胞中过表达可增强 TRM核心标志物表达。本研究轨迹分析提示,KLF6 参与随年龄增长获得成熟成人样 TRM表型的过程,为儿童期 TRM阶段性成熟提供潜在机制。 本研究结果表明,人类记忆 T 细胞早期发育过程中优先启动转录调控程序(尤其黏膜部位)。尽管抗原特异性尚不明确,但其时间窗与共生微生物定植、经呼吸 / 摄入接触黏膜病原体的动态过程一致。在这一抗原暴露密集的关键阶段,限制促炎应答与组织损伤风险具有重要意义。HELIOS 上调可作为 Treg介导外周耐受的补充机制。 随年龄增长的组织驻留程序诱导,也可能源于组织自身微环境、抗原暴露或其他组织因子的年龄相关变化。环境因素(微生物暴露、感染史、疫苗接种)如何塑造生命早期的这些转录程序,仍有待进一步探究。 综上,本研究鉴定的年龄特异性转录因子,揭示了调控生命早期免疫力的潜在靶点,为优化婴幼儿疫苗与免疫治疗策略提供理论基础。 四、方法(精简) 4.1 人类器官供体与组织获取 经美国器官获取组织批准的研究方案,获取离世(脑死亡)器官捐献者的淋巴与黏膜组织;成人组织来自纽约大都会区 LiveOnNY,婴儿组织来自 LiveOnNY 及新生儿发育人类图谱计划(HANDEL)。所有供体(n=32,年龄 0.1–78 岁,男性 26 例、女性 6 例)无癌症、慢性疾病,乙肝 / 丙肝 / HIV 阴性,无活动性感染证据。哥伦比亚大学伦理委员会确认,离世器官供体组织研究不属于人类受试者研究。 4.2 组织处理、T 细胞分离与刺激 组织冰上转运,2–24 小时内处理为单细胞悬液;密度梯度离心分离单核细胞,磁珠阴性分选富集 T 细胞(纯度 > 80%–95%)。分静息组(完全培养基培养 16 小时)与活化组(CD3+CD28 抗体刺激 16 小时),去除死细胞后进行单细胞包裹。 4.3 scRNA‑seq 与数据分析 采用 10x Genomics 平台建库测序,STAR 比对参考基因组,EmptyDrops 去除背景 RNA,GMM 过滤低质量细胞;Phenograph 无监督聚类,朴素贝叶斯分类器划分 T 细胞亚群;consensus‑scHPF 解析转录因子,ARACNe 重构调控网络,VIPER 分析转录因子活性,GSEA 关联转录因子与 scHPF 因子。 4.4 snRNA‑seq/ATAC‑seq 联合分析 分离脾脏 T 细胞细胞核,10x Genomics Multiome 平台建库测序;Cell Ranger 比对,muon 与 Scanpy 质控、整合、聚类;Diffusion map 构建拟时轨迹,MACS3 峰检出,chromVAR 分析转录因子基序可及性。 4.5 流式细胞术与细胞内染色 表面标志物与转录因子染色,PMA + 离子霉素刺激检测细胞因子分泌,Cytek Aurora 光谱流式获取数据,FlowJo 分析。 4.6 CRISPR–Cas9 敲除 Cas9 核糖核蛋白转染原代婴儿 T 细胞,靶向 IKZF2;培养 3 天后流式检测 HELIOS 表达,scRNA‑seq 分析转录变化。 4.7 统计学分析 GraphPad Prism 统计,双侧 t 检验;多元线性回归关联 scHPF 因子与协变量;FDR 校正多重检验,P<0.05 为差异显著。More
不同转录因子调控婴儿与成人记忆 T 细胞的组织适应性及效应功能 期刊:Nature Immunology(2026) 发表时间:2026 年 6 月 2 日 DOI:10.1038/s41590-026-02535-1 摘要 生命早期是建立记忆 T 细胞库的关键阶段,记忆 T 细胞在婴儿期可快速定植于黏膜组织,但这些新生记忆 T 细胞的免疫保护能力弱于成人。本研究对婴儿(2–9 月龄)与成人(40–63 岁)器官供体的淋巴组织与黏膜组织中静息及 CD3+CD28 抗体活化的 T 细胞进行单细胞 RNA 测序,探究人类记忆 T 细胞功能调控的年龄依赖性机制。 结果显示,婴儿CCL5 + 效应记忆 T 细胞的效应功能弱于成人。转录因子调控网络分析鉴定出HELIOS与KLF6分别为婴儿与成人组织记忆 T 细胞状态的核心调控因子。结合单细胞核 RNA 测序、转座酶可及染色质测序与 CRISPR–Cas9 敲除实验,证实HELIOS(IKZF2)是婴儿 CCL5 + 效应记忆 T 细胞特异性转录程序的关键调控因子,可抑制 SELL+CCR7 + 初始 T 细胞 / 中央记忆 T 细胞的效应功能。本研究揭示了生命早期调控 T 细胞功能状态的核心分子机制。 正文 一、引言 生命早期适应性免疫系统的成熟对建立终身免疫记忆与防御能力至关重要。婴幼儿阶段会持续接触共生菌、致病菌、疫苗及环境抗原,触发强烈的初次免疫应答并形成记忆 T 细胞,介导后续的回忆反应。但相较于成人,婴儿对呼吸道、胃肠道等常见病原体高度易感,提示其记忆应答发育尚不完善。 小鼠研究已证实,组织驻留记忆 T 细胞(TRM)介导黏膜与屏障部位的免疫保护;但人类生命早期组织记忆应答的功能与建立机制仍不明确,而这对优化婴幼儿疫苗策略与免疫治疗至关重要。 婴儿小鼠与人类的 T 细胞以初始 T 细胞(TN)为主。婴儿小鼠感染模型显示,婴儿 TN可分化为效应 T 细胞(Teff),但针对全身性与呼吸道病毒感染的循环及组织定位记忆 T 细胞生成量更少、保护力更弱;婴儿疫苗诱导的记忆 T 细胞数量与保护力也显著低于成年小鼠。 婴儿与成人 T 细胞的原发性应答差异源于细胞固有机制,包括 T 细胞受体(TCR)敏感性、造血前体细胞特性及胸腺新近输出细胞的影响。然而,婴儿记忆 T 细胞应答欠佳的分子机制在小鼠中尚未阐明,在人类中更是完全未知。 借助人类跨年龄段器官供体的多组织样本,研究者已明确人类组织驻留记忆 T 细胞(TRM)的核心特征、黏膜组织分区、位点特异性适应及年龄稳定性。出生后 1–2 年内,具有经典 CD45RA−CCR7−效应记忆 T 细胞(TEM)表型并表达 CD69(TRM标志性分子)的 T 细胞迅速定植于黏膜组织,但这类细胞的核心及组织特异性 TRM标志物表达水平更低,且产生促炎细胞因子(IFN-γ、TNF)的能力弱于成人及年长儿童的黏膜 TRM。目前尚不清楚调控婴儿记忆 T 细胞独特表型的转录机制,以及组织适应性黏膜记忆 T 细胞的成熟通路。 本研究采用单细胞 RNA 测序(scRNA‑seq)分析婴儿与成人淋巴组织、黏膜组织来源的 T 细胞(含 CD3+CD28 抗体活化组),应用一致性单层级泊松因子分解(consensus‑scHPF)解析婴儿与成人 T 细胞活化、效应功能及组织适应相关的转录状态。 研究发现,婴儿组织记忆 T 细胞呈现干细胞样转录特征(TCF1、LEF1、SOX4),但对 TCR 介导刺激的转录应答弱于成人。通过概率建模与调控网络推断,鉴定出HELIOS(IKZF2)与KLF6为年龄特异性转录因子,分别调控婴儿与成人 T 细胞的功能与组织适应。利用 CRISPR–Cas9 敲除、单细胞核 RNA 测序(snRNA‑seq)与转座酶可及染色质测序(ATAC‑seq)证实,HELIOS 是婴儿记忆 T 细胞的关键调控因子,可维持干细胞样状态、抑制效应分化及组织特征表达。综上,本研究揭示年龄调控记忆 T 细胞命运与功能的核心机制,为生命早期 T 细胞应答靶向干预提供理论依据。 二、结果 2.1 单细胞 RNA 测序绘制不同年龄与组织的 T 细胞亚群图谱 为明确生命早期不同组织 T 细胞的转录编程,本研究对婴儿(n=4,2–9 月龄,男女各 2 例)与成人(n=4,40–63 岁,男性 4 例)的淋巴组织、黏膜组织 T 细胞进行 scRNA‑seq 检测。样本取自离世器官捐献者,包括淋巴器官(骨髓、脾脏、扁桃体、肠道派尔集合淋巴结、肺 / 空肠 / 结肠相关淋巴结)与黏膜 / 屏障组织(肺、空肠、回肠、结肠)。 经磁珠阴性分选富集 T 细胞,分为静息组(培养基培养)与活化组(CD3+CD28 抗体刺激),采用 10x Genomics Chromium 平台进行单细胞测序。数据集与已发表的成人器官供体及健康献血者静息 / 活化 T 细胞数据合并,最终获得约 27.5 万个跨 12 种组织的 T 细胞单细胞图谱。 合并数据集采用朴素贝叶斯分类器划分 T 细胞亚群:SELL+CCR7 + 初始 T 细胞 / 中央记忆 T 细胞(TN/TCM)、CCL5 + 效应记忆 T 细胞(TEM,含 CD4 + 与 CD8+)、CD4+FOXP3 + 调节性 T 细胞(Treg)及 γδ T 细胞。均匀流形逼近与投影(UMAP)显示,年龄与刺激条件是导致转录组变异的主要因素。婴儿与成人各 T 细胞亚群的经典标志物表达高度保守。 为对比婴儿与成人跨组织 T 细胞谱,聚焦两组共有的组织样本:血液、骨髓、肺 / 空肠相关淋巴结、脾脏、肺、空肠、结肠。婴儿血液与淋巴部位的 T 细胞以 SELL+CCR7+ TN/TCM为主,仅少量 CD4+FOXP3+ Treg及 CCL5+ TEM/γδ T 细胞;婴儿黏膜组织与脾脏中 CCL5+ TEM比例更高,尤以 CD8+CCL5+ TEM显著,肠道 T 细胞几乎均为 CCL5+ TEM,与既往报道一致。 与之相反,成人黏膜组织与脾脏中 CCL5+ TEM占绝对优势,比例显著高于 SELL+CCR7+ TN/TCM;成人多数组织中 CD8+ SELL+CCR7+ TN/TCM比例远低于 CD4 + 亚群。上述结果表明,婴儿与成人 T 细胞亚群组成存在组织特异性年龄差异,与既往流式细胞术结果相符。 2.2 婴儿效应记忆 T 细胞对 TCR 刺激呈低反应性 婴儿与成人 T 细胞亚群的差异表达分析显示,CCL5+ TEM在静息与活化状态下均存在显著年龄差异。静息状态下,婴儿 CCL5+ TEM高表达干细胞样记忆相关转录因子TCF7(TCF1)、LEF1、KLF2,以及 Wnt 信号通路 TCF‑LEF 家族辅因子SOX4与成人 Treg特征性分子IKZF2(HELIOS)。 婴儿 CD4+CCL5+ TEM富集 Th2 驱动转录因子GATA3,CD8 + 亚群高表达组织驻留相关ZNF683(Hobit)与干细胞样记忆前体标志物ID3。而成人 CCL5+ TEM偏好表达组织适应与黏附相关基因,如LGALS1、LGALS3、ANXA1、VIM、ITGA1。 流式细胞术证实,婴儿脾脏 CD8+CCR7−TEM中 TCF1 与 LEF1 表达显著高于成人,CD4+CCR7−TEM中仅 LEF1 显著升高。 差异表达分析发现,成人 CD3+CD28 抗体刺激 T 细胞(尤其 CCL5+ TEM)中促炎细胞因子、趋化因子与细胞毒性介质(IFNG、IL‑2、TNF、CCL3、CCL4、GZMB、PRF1)的转录本诱导更强。流式细胞术显示,成人脾脏 CD4+CCR7−TEM与 CD8+CCR7−TEM中 IFNγ、IL‑2、TNF 阳性细胞比例显著高于婴儿。 综上,婴儿 CCL5+ TEM呈现独特转录状态:干性相关转录因子高表达,效应分子表达低于成人 CCL5+ TEM。 2.3 T 细胞呈现年龄与组织特异性表达程序 为揭示婴儿与成人组织 T 细胞的独特基因表达程序,采用consensus‑scHPF(贝叶斯因子分解法)挖掘单细胞中共同表达的潜在转录特征(因子),并以年龄、组织定位、亚群、谱系、活化状态为协变量进行多元线性回归,关联各因子与临床表型。 分析鉴定出 18 种独立特征,对应 T 细胞亚群、代谢、组织适应、活化状态及 / 或效应功能。 consensus‑scHPF 揭示 3 种与组织定位及适应相关、且婴儿与成人存在差异的因子: 婴儿组织因子:在婴儿 CCL5+ TEM中高差异表达,含 SMC4、NCR3、CXXC5、LAYN、IKZF2,各组织中婴儿 CCL5+ TEM均显著富集。 组织特征因子:在成人 CD4+/CD8+CCL5+ TEM中富集,含 S100A4/6、CRIP1、LGALS1、KLRB1、ANXA2。 肠道驻留因子:以肠道归巢与黏附标志物(CCR9、ITGA1、CTNNA1)及组织驻留记忆 T 细胞发育相关因子(AHR、JUN、FOSB)为特征,显著偏向成人 CD4+/CD8+CCL5+ TEM(尤其黏膜组织)。 另有 3 种与效应状态相关的因子:细胞毒性因子(GZMK、GNLY、GZMA、NKG7、PRF1)、趋化 / 细胞毒性因子(GZMB、GZMH、FASLG、PRF1、CCL3、CCL4)、炎性细胞因子因子(IFNG、IL‑2、TNF、CSF2、LTA)。三者均在成人 CCL5+ TEM中富集,且后两者在活化 CCL5+ TEM中高于静息组。 整体表明,婴儿与成人 T 细胞的组织适应程序及TCR 刺激应答存在显著差异。 对 CD3+CD28 抗体刺激后 CCL5+ TEM的分析显示,成人活化 CCL5+ TEM(尤其 CD8 + 亚群)中炎性细胞因子与趋化 / 细胞毒性因子的核心基因表达更高。扩散映射活化轨迹显示,成人 CD8+CCL5+ TEM沿活化轴的细胞数量远多于婴儿,提示婴儿 T 细胞刺激后的转录应答减弱、效应功能受限。 在 3 种组织驻留 / 适应相关因子中,婴儿组织因子核心基因高度富集于婴儿 CD4+/CD8+CCL5+ TEM;组织特征因子在婴儿与成人 CCL5+ TEM中广泛表达;肠道驻留因子仅富集于成人肠道部位(空肠、结肠、空肠相关淋巴结)的 CCL5+ TEM。空肠静息 CCL5+ TEM的成熟轨迹显示,婴儿组织因子与肠道驻留因子分别特异性富集于婴儿与成人细胞,而组织特征因子为两者共有,提示其代表空肠组织适应的中间状态。 2.4 T 细胞激活年龄特异性转录因子 为鉴定执行组织与年龄特异性表达程序的转录因子,应用ARACNe 算法重构基因调控网络,生成各转录因子的靶基因集(调控子);再通过基因集富集分析(GSEA)将转录因子与 scHPF 因子定义的细胞状态关联。 该分析鉴定出多种已知转录因子‑T 细胞状态关系:IRF1 与 STAT1 调控干扰素信号应答、NFKB1 调控活化、IRF7 调控 Treg干扰素应答、ZEB2 调控细胞毒性 T 细胞功能、KLF2/TCF1/LEF1 调控初始 T 细胞干性与静息态。同时发现,HELIOS(IKZF2)是婴儿组织因子的首要调控因子,KLF6是组织特征因子与肠道驻留因子的首要调控因子。 进一步通过VIPER 分析在单细胞水平推断转录因子活性。空肠组织适应轨迹的映射结果证实:HELIOS 活性仅存在于婴儿,KLF6 活性随婴儿向成人过渡而增强,并在所有成人 CD4+/CD8+CCL5+ TEM中升高。 组织驻留相关转录因子中,RUNX3 在婴儿与成人 CCL5+ TEM均有活性,AHR 活性显著富集于成人 CCL5+ TEM;干性转录因子 LEF1、TCF1 活性显著富集于婴儿 CCL5+ TEM。LEF1 调控子与组织特征因子、肠道驻留因子核心基因(含 KLF6)存在大量抑制性相互作用,提示 LEF1 可能拮抗组织适应所需的转录程序。 综上,本分析发现与 T 细胞状态相关的关键转录因子,并确定HELIOS是 T 细胞中婴儿特异性组织适应程序的候选调控因子。 2.5 HELIOS 调控婴儿 T 细胞的效应功能 为探究 HELIOS 在婴儿 CD8+ TEM中的表达与活性调控,对额外 3 例婴儿(0.25–1 岁,男性)与 2 例成人(45–51 岁,男性)供体的脾细胞进行snRNA‑seq/ATAC‑seq 联合测序。通过婴儿与成人 T 细胞的染色质可及性差异,鉴定年龄依赖性基因表达的顺式调控元件,并以差异可及染色质峰中的转录因子基序富集作为年龄依赖性转录因子活性的正交验证。 整合 snRNA‑seq/ATAC‑seq 的 CD8+CCL5+ TEM图谱,鉴定出 1 种婴儿特异性细胞状态与 2 种成人细胞状态。构建基于 mRNA 的拟时轨迹,解析从婴儿状态向两种成人状态过渡过程中的基因表达与转录因子活性变化。 第一种成人 CD8+CCL5+ TEM状态:细胞毒性与活化标志物(GZMA、GZMH、GNLY、CCL4、CCL5)高于婴儿状态; 第二种成人 CD8+CCL5+ TEM状态:共刺激标志物(CD28、ICOS)与转录因子 BATF 高表达。 结合同一细胞核的基因表达与染色质可及性数据,进行转录因子基序富集分析,并沿婴儿→成人拟时轨迹映射转录因子活性。结果显示,HELIOS 活性在两种轨迹中均显著富集于婴儿 CD8+CCL5+ TEM状态,在独立器官供体队列中从染色质水平独立验证了 scRNA‑seq 与调控网络的发现。 利用 CD8+CCL5+ TEM的 snATAC‑seq 数据,评估 IKZF2 表达是否受年龄依赖性顺式调控。在 IKZF2 位点上游鉴定出 1 个顺式调控元件,其染色质可及性峰在婴儿 CD8+CCL5+ TEM中富集;该区域含多个婴儿特异性活性转录因子(SOX4、ETV1、TCF7L1)的结合基序,以及人群遗传研究中与免疫细胞 IKZF2 表达相关的表达数量性状位点(eQTLs),提示 IKZF2 表达通过该顺式调控元件受成人 CD8+CCL5+ TEM中活性减弱的转录因子调控。 流式细胞术显示,婴儿 T 细胞中 HELIOS 表达显著高于成人,涵盖 CD8+CD45RA+CCR7+ TN、CD8+CD45RA−CCR7+ TCM、CD8+CCR7−TEM,CD4+CD45RA+CCR7+ TN中差异稍弱。婴儿与成人的 CD8+/CD4+ T 细胞经 CD3+CD28 抗体刺激后,HELIOS 的 mRNA 与蛋白表达均未上调。 采用CRISPR–Cas9 基因编辑敲除原代婴儿 T 细胞中的 HELIOS。对 2 例婴儿供体(2–3 月龄,男性)脾脏分离的 CD3+ T 细胞,转染靶向 IKZF2 的 Cas9 核糖核蛋白(未预先刺激 T 细胞),流式细胞术检测敲除效率。结果显示,HELIOS 敲除使 γδTCR−FOXP3−CD3+ T 细胞中 HELIOS + 细胞比例降低约 60%。 对静息状态下婴儿 HELIOS 敲除与野生型 CD3+ T 细胞的 scRNA‑seq 分析显示,敲除组 CD8+CCL5+ TEM中 HELIOS 激活靶基因(LAYN、PDE7B、CHST2、SMC4、CXXC5)表达下调,CRISPR–Cas9 敲除验证了计算推断的 HELIOS 转录靶标。 HELIOS 敲除与野生型 T 细胞的大部分转录差异出现在 CD3+CD28 抗体刺激的 CD4+SELL+CCR7+ TN–TCM与 CD8+SELL+CCR7+ TN–TCM中,包括促炎细胞因子、趋化因子(IL‑2、CSF2、XCL1、XCL2、CCL3、CCL4、CCL20)与共刺激分子(TIGIT、KLRB1、TNFRSF4、TNFRSF9、TNFRSF18、TNFSF14、FCER1G、IL18R1、IL1R1)转录本表达升高,提示TCR 介导的活化增强。 4 小时 PMA + 离子霉素刺激后,婴儿 HELIOS 敲除组 CD4+CD45RA−CCR7+ TCM与 CD4+CCR7−TEM中 IL‑2 + 细胞比例高于野生型。scHPF 分析显示,与 T 细胞活化及效应功能相关的趋化 / 细胞毒性因子、炎性细胞因子因子,在 HELIOS 敲除婴儿 T 细胞(尤其 CD4+/CD8+SELL+CCR7+ TN–TCM)中显著富集,表明HELIOS 缺失使婴儿 T 细胞对刺激的转录应答更接近成人。 静息状态下 HELIOS 敲除 T 细胞的转录因子分析显示,KLF6及其他 T 细胞效应功能调控因子(IRF1、IRF2、STAT5A、IKZF3、RORA)表达升高,多数与成人 SELL+CCR7+ TN–TCM相关。CD3+CD28 抗体刺激后,婴儿 HELIOS 敲除 SELL+CCR7+ TN–TCM中干性转录因子 TCF7、LEF1 表达降低(二者为婴儿野生型 SELL+CCR7+ TN–TCM的特征)。 综上数据表明,HELIOS 缺失驱动婴儿 T 细胞从婴儿样转录状态向成人样状态转变。 三、讨论 本研究系统分析了人类组织中婴儿与成人 T 细胞的功能应答及组织定位,发现婴儿期记忆 T 细胞主要局限于黏膜部位,功能呈低反应性,组织驻留关键分子表达低于成人记忆 T 细胞;同时鉴定出HELIOS与KLF6分别为调控婴儿与成人组织记忆 T 细胞功能及组织驻留的核心转录因子。 snATAC‑seq 证实 HELIOS 调控活性特异性富集于婴儿记忆 T 细胞;婴儿 T 细胞中 HELIOS 敲除可增强活化与效应功能,使其趋近成人记忆 T 细胞。本研究揭示了控制组织适应与效应应答的年龄特异性转录程序,作为生命早期限制记忆 T 细胞功能的核心机制。 通过 scRNA‑seq 对比黏膜与淋巴部位静息、活化状态下婴儿与成人 T 细胞的转录谱,从多方面证实婴儿 TEM处于分化程度更低、功能未成熟的状态: 婴儿 TEM即使在刺激后仍维持干性标志物(SOX4、TCF7、LEF1)表达,成人 TEM则不表达; 婴儿 TEM的组织驻留标志物(ITGA1、ITGAE、CXCR6)与位点特异性适应标志物(如 CCR9)表达低于成人,与既往报道一致; 既往研究显示婴儿 TEM经 PMA + 离子霉素非特异性活化后,效应细胞因子分泌少于成人 TEM;本研究进一步发现,婴儿记忆 T 细胞对TCR 偶联刺激基本无反应,不具备成人 TEM活化标志性的快速回忆应答,提示存在婴儿 T 细胞固有程序介导的耐受状态。 本研究将HELIOS鉴定为婴儿 T 细胞活化与分化的核心转录调控因子与抑制因子。婴儿 T 细胞中 HELIOS 转录因子基序活性显著高于成人;特异性敲除 HELIOS 可增强婴儿 T 细胞的活化与效应功能。值得注意的是,静息婴儿 T 细胞中敲除 HELIOS 可诱导成人样转录因子(含 KLF6)表达,CD3+CD28 抗体刺激后高表达 CCL3、CCL4、IL‑2 等趋化因子与细胞因子基因,趋近成人组织记忆 T 细胞。 HELIOS 最初被发现表达于小鼠与人 Treg,可沉默 IL2 位点;IKZF2 功能缺失与错义突变患者的 T 细胞呈现 IL‑2 分泌增强或异常。外周血新生儿与胎儿 T 细胞(以初始表型为主)的 HELIOS 表达也高于成人。本研究发现,婴儿初始 T 细胞的 IKZF2 表达高于成人,且婴儿 TEM的 IKZF2 表达显著高于初始 T 细胞,据此提出HELIOS 作为组织定位 TEM中促炎应答与驻留的细胞内源性抑制因子。 通过基因调控网络推断,将KLF6鉴定为跨多组织的组织特征及肠道特异性驻留特征的候选转录调控因子,与成人空肠 T 细胞的肠道驻留特征相似。小鼠研究中 Klf6 被鉴定为 TRM特异性转录因子,在抗原特异性 T 细胞中过表达可增强 TRM核心标志物表达。本研究轨迹分析提示,KLF6 参与随年龄增长获得成熟成人样 TRM表型的过程,为儿童期 TRM阶段性成熟提供潜在机制。 本研究结果表明,人类记忆 T 细胞早期发育过程中优先启动转录调控程序(尤其黏膜部位)。尽管抗原特异性尚不明确,但其时间窗与共生微生物定植、经呼吸 / 摄入接触黏膜病原体的动态过程一致。在这一抗原暴露密集的关键阶段,限制促炎应答与组织损伤风险具有重要意义。HELIOS 上调可作为 Treg介导外周耐受的补充机制。 随年龄增长的组织驻留程序诱导,也可能源于组织自身微环境、抗原暴露或其他组织因子的年龄相关变化。环境因素(微生物暴露、感染史、疫苗接种)如何塑造生命早期的这些转录程序,仍有待进一步探究。 综上,本研究鉴定的年龄特异性转录因子,揭示了调控生命早期免疫力的潜在靶点,为优化婴幼儿疫苗与免疫治疗策略提供理论基础。 四、方法(精简) 4.1 人类器官供体与组织获取 经美国器官获取组织批准的研究方案,获取离世(脑死亡)器官捐献者的淋巴与黏膜组织;成人组织来自纽约大都会区 LiveOnNY,婴儿组织来自 LiveOnNY 及新生儿发育人类图谱计划(HANDEL)。所有供体(n=32,年龄 0.1–78 岁,男性 26 例、女性 6 例)无癌症、慢性疾病,乙肝 / 丙肝 / HIV 阴性,无活动性感染证据。哥伦比亚大学伦理委员会确认,离世器官供体组织研究不属于人类受试者研究。 4.2 组织处理、T 细胞分离与刺激 组织冰上转运,2–24 小时内处理为单细胞悬液;密度梯度离心分离单核细胞,磁珠阴性分选富集 T 细胞(纯度 > 80%–95%)。分静息组(完全培养基培养 16 小时)与活化组(CD3+CD28 抗体刺激 16 小时),去除死细胞后进行单细胞包裹。 4.3 scRNA‑seq 与数据分析 采用 10x Genomics 平台建库测序,STAR 比对参考基因组,EmptyDrops 去除背景 RNA,GMM 过滤低质量细胞;Phenograph 无监督聚类,朴素贝叶斯分类器划分 T 细胞亚群;consensus‑scHPF 解析转录因子,ARACNe 重构调控网络,VIPER 分析转录因子活性,GSEA 关联转录因子与 scHPF 因子。 4.4 snRNA‑seq/ATAC‑seq 联合分析 分离脾脏 T 细胞细胞核,10x Genomics Multiome 平台建库测序;Cell Ranger 比对,muon 与 Scanpy 质控、整合、聚类;Diffusion map 构建拟时轨迹,MACS3 峰检出,chromVAR 分析转录因子基序可及性。 4.5 流式细胞术与细胞内染色 表面标志物与转录因子染色,PMA + 离子霉素刺激检测细胞因子分泌,Cytek Aurora 光谱流式获取数据,FlowJo 分析。 4.6 CRISPR–Cas9 敲除 Cas9 核糖核蛋白转染原代婴儿 T 细胞,靶向 IKZF2;培养 3 天后流式检测 HELIOS 表达,scRNA‑seq 分析转录变化。 4.7 统计学分析 GraphPad Prism 统计,双侧 t 检验;多元线性回归关联 scHPF 因子与协变量;FDR 校正多重检验,P<0.05 为差异显著。More